Newport Laboratory: Fisheries Behavioral Ecology Program
Density-Dependent Habitat Selection In Juvenile Flatfishes
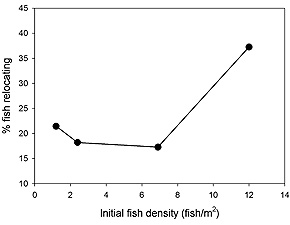
Figure 1. Net rates of redistribution of northern rock sole by nighttime water column swimming as a function
of fish density. The rate of nighttime redistribution was not affected by the presence of benthic structure (sponges)
in either side of the experimental arenas. |
|
The distribution of juvenile fishes in nursery grounds is determined by behavioral responses to environment variables
(sediment type, structure, prey availability). Therefore, the distributions are assumed to reflect habitat preference and
used to determine which habitats are deemed “essential” or “critical” to the population. However, the relationships between
fish distribution and habitats are complicated by density-dependent responses.
The reasons for density-dependent responses are not known but are generally assumed to be related to competition for food
or conspicuousness to predators. As a result, habitats that are not preferred when populations are at low densities are
used to a greater degree when density in the preferred habitat rises. The principles of density-dependent habitat
selection were first outlined for birds, but these models have subsequently been adapted to fisheries science with the
“basin hypothesis.”
In general, these models predict how motile animals should distribute themselves as high-quality habitats become saturated
with conspecifics. The effects of density on habitat use by juvenile flatfishes are the subject of several related projects
being conducted in the Fisheries Behavioral Ecology Program (FBEP).
In one series of experiments, Thomas Hurst and intern Clara Lampi examined how density of northern rock sole affected the
frequency of nighttime swimming behavior. In this experiment, fish were initially stocked onto one side of a 3-m diameter
tank at densities from 1.2 to 12 fish m-2. The fish were allowed to swim over a barrier to move to the other side of the
tank which was not stocked with fish.
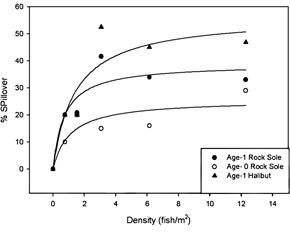
Figure 2. Percentage spillover (fraction of fish) from a preferred substrate (sand) to the non-preferred substrate
(gravel) as a function of fish density. Data is for age-0 northern rock sole, age-1 northern rock sole and age-1
Pacific halibut. |
|
Nighttime swimming of flatfishes in the water column and at the surface is a widely observed but poorly understood aspect of
the behavior of juvenile flatfishes. The use of nighttime swimming by northern rock sole was described in a recent paper
(Hurst and Duffy 2005) and is believed to be an important mechanism of redistribution in heterogeneous environments.
Per-capita rates of redistribution (into empty habitat) increased from approximately 18% at the lower stocking densities to
30% at the highest stocking density (Fig. 1 above). Additional trials indicated that nighttime redistribution was not
sensitive to the presence of epibenthic structure. Hence, structured habitats that fish congregate in during the day may
“leak” fish into surrounding habitats at night when densities reach certain levels.
In separate experiments, Ben Laurel is comparing the density-dependent habitat selection of juvenile northern rock sole
and Pacific halibut. Both species are known to strongly prefer sandy sediment over coarser gravels. In this experiment, the
preference of fish for sand substrate is tested at varying fish densities and sizes to determine if increased fish densities
result in greater usage of the less preferred substrate. Preliminary results (Fig. 2 above) indicate that habitat selection
of age-0 rock sole is less sensitive to fish density than that of age-1 rock sole or age-1 halibut. Age-0 rock sole were less
likely to leave the preferred substrate as fish density increased.
| |

Figure 3. Photograph of camera sled developed by FBEP researchers and used to examine the habitat characteristics
and distribution of juvenile flatfishes in nearshore Kodiak Island nursery areas. |
Examination of the interaction between fish density and habitat selection is also being conducted in the field. FBEP
researchers have developed a towed camera sled for the examination of juvenile flatfish distributions and habitat
associations (Fig. 3). The benefit of this camera system is that it provides continuous and spatially-explicit (through
navigation linkage) sampling of fish, benthic bedform, water temperature, and biological components of habitat not
effectively sampled with trawls. This camera system has been used to survey flatfishes and their habitats in several Kodiak
Island nursery areas over the last 4 years. Data from these surveys, analyzed at multiple spatial scales with generalized
additive models (GAM), demonstrate a strong nonlinear effect of worm tubes and other structural features on flatfish density,
in addition to effects of sediment type and bedform.
In an extension of this project, Ben Laurel is using data on the fine-scale distribution of juvenile flatfishes in Kodiak
Island nursery areas to examine the patchiness of fish and habitats. Patchiness measures are often used to describe the
variable spatial and temporal patterns in the distribution of marine fish. Such measures are particularly useful during early
life stages as they describe the points where passive contributions (i.e., larval drift) weaken and behavioral processes
begin. Although several studies have measured patchiness in the field, none have explicitly linked active habitat selection
with patchiness estimates. Successfully linking patchiness measures to process-derived measures of habitat selectivity will
provide a powerful tool in categorizing essential fish habitat for exploited marine species.
By Thomas Hurst
 |

|
OND2005 quarterly sidebar
AFSC Quarterly
Research Reports Oct-Dec 2005
Contents
Feature
ABL Reports
FMA Reports
NMML Reports
RACE Reports
REFM Reports
Milestones
Quarterly Index
Quarterly Home
|
