Bering Sea/Aleutian Island (BSAI) region Tanner crab, classified as a Tier 4 stock, are characterized by estimates of biomass, but with little habitat or ecological information guiding assessment. Tanner stocks (Gulf of Alaska and Bering Sea) are depressed or rebuilding. Stock declines are little understood but generally are attributed to over-fishing or climatic changes. Essential fish habitat knowledge for juvenile Tanner crab (age-0) is rudimentary at best. Our 2010 research examined habitat use by age-0 Tanner crabs in four Kodiak embayments: Pillar Creek Cove, Holiday Beach, Kalsin Bay, and Womens Bay (Fig. 1). Womens Bay, in particular, was chosen because it supports a Tanner crab subsistence fishery and is regarded as a crab nursery. By the end of summer 2010, age-0 Tanner crabs from Womens and Kalsin Bays were one molt stage larger than those in the Pillar and Holiday sites. Larger size at the end of the first summer could provide increased fitness to Womens Bay crabs, as they are less vulnerable to predation. Further, since Womens Bay crabs are one stage larger at summers end, they will likely reach reproductive age 1 year sooner. Understanding how nursery/habitat influences growth and size/age at maturity provides a direct linkage between essential fish habitat and crab stocks. Following on from 2010, our 2011 field work focused on several specific hypotheses addressing how these juvenile Tanner nurseries may be functioning.
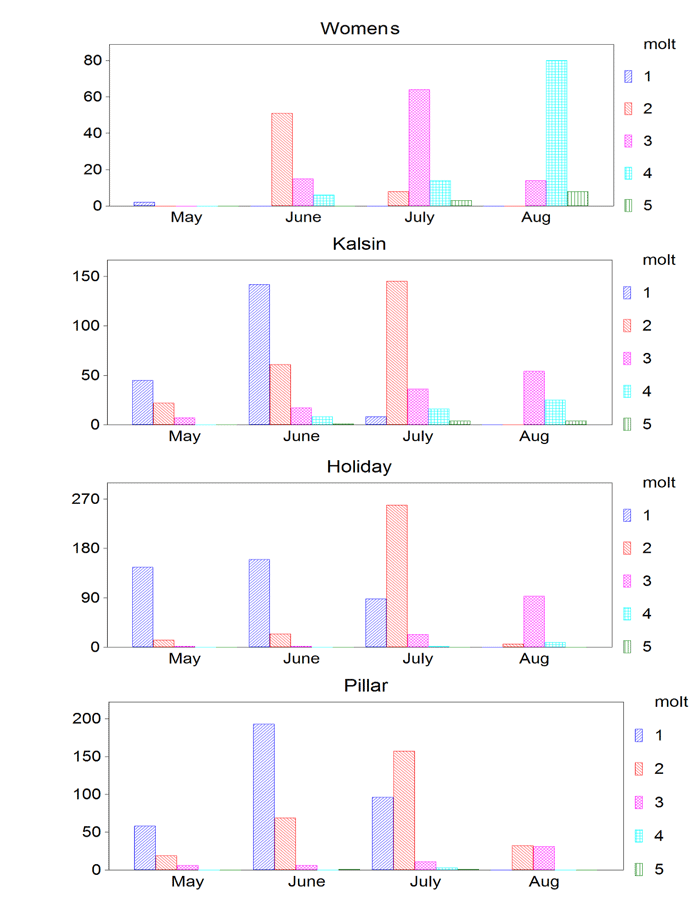
Figure 2. Relative size frequency distributions of age-0 yr Tanner crabs from each site, May – August. Click here to enlarge.
Of potential causative factors contributing to the larger size of juvenile Tanner crabs from Womens and Kalsin Bays, the first factor to consider involves possible differences in the timing of recruitment; crabs may recruit earlier to Womens and Kalsin, compared to Holiday and Pillar. To examine this, we compared the May 2011 size-frequency distributions in all four embayments (Fig. 2, G = 32.57, df = 12, P = 0.001). The major difference involved Womens, where age-0 crabs (C1-C3) were nearly absent in May. Yet by mid-June, crabs appearing at Womens were already C2 or larger; one molt more than crabs at the other sites. Crabs in Kalsin were more comparable to those in Holiday and Pillar. This molt advantage attained by crabs from Womens Bay in June persisted throughout the summer, which leads us to conclude that early recruitment cannot explain the larger crabs at Womens Bay.
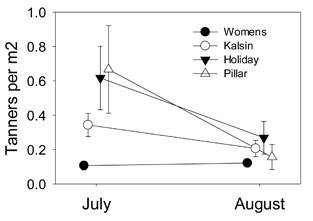
A second factor to consider involves potential difference in size-selective predation between embayments. Perhaps there is strong predation upon smaller crabs in Womens Bay, leading to a shifted size-frequency distribution and the “appearance” of more rapid growth. This should also be manifest by an overall larger change in population size during the post recruitment period. Recruitment was largely complete by July 2011 (Fig. 2) and, therefore, population changes from July to August were due to mortality or emigration/immigration. Neither Womens nor Kalsin Bays experienced significant population declines from July to August (Fig. 3, Womens: Wilcoxon, P = 0.413, Kalsin: P = 0.204). At Pillar, crab abundances declined significantly (Wilcoxon U, P = 0.040). The decline at Holiday was not significant (Wilcoxon, P = 0.419). Crabs were surviving and remaining resident at Womens and Kalsin Bays, and to a lesser extent at Holiday, but dying and/or emigrating at Pillar. Therefore, we can reject the size- selective predation hypothesis as an explanation for larger crabs at Womens Bay.
Figure 3. July to August population size changes at each site.
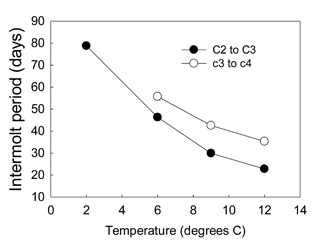
Figure 4. Influence of temperature on C2-C3 and C3-C4 inter-molt periods.
It may be that crabs are indeed growing faster at Womens Bay. Water temperature is often a critical factor influencing the growth of juvenile crabs and fishes. Therefore, we conducted a laboratory experiment to examine temperature dependence of growth in juvenile Tanner crabs. Higher temperature significantly accelerated Tanner growth in the lab (Fig. 4). The C2-C3 intermolt period was 46 days at 6°C, but only 30 days at 9°C. Similarly, the C3-C4 intermolt period was 56 days at 6°C but 43 days at 9°C. Yet temperature loggers at each site (~15m) during 2001 indicated only minor temperature differences. During the early summer, when crabs at Womens Bay appeared to undergo accelerated growth, the average difference in temperature between sites was only 0.3°C. Therefore, we consider it unlikely that temperature, at the depths we studied, was a significant contributor to the differential growth between populations.
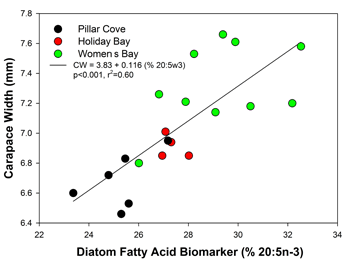
A final factor we considered involved food quality. More specifically, we hypothesized that food quality may be better at Womens Bay, leading to more rapid growth. We reasoned that this should be reflected in the condition and tissues of crabs from the different embayments. In 2011, crabs were collected for extraction and analysis of storage lipids and fatty acid biomarkers. Unfortunately, these samples were lost in a freezer malfunction (word to the wise, check alarm systems). However, samples taken at Womens, Pillar, and Holiday during July 2010 suggest a possible difference in diet between sites. Significant differences occurred in the amount of storage lipid in crabs from the three coves. Pillar crabs had significantly lower storage lipid (triacylglyercols, ~8%) compared to Holiday and Womens (~20%). Womens crabs had consistently higher levels of fresh diatom fatty acid biomarkers than crabs from the other embayments. More specifically, the levels of 20:5n-3 was significantly higher in crabs from Womens. This specific essential fatty acid has previously been demonstrated to accelerate growth in larval Pacific cod, larval rock sole, and wild red king crab juveniles. Not only was this essential fatty acid and diatom marker found at higher levels in Womens Bay crabs, it was strongly correlated with the carapace widths of juvenile C3 crabs across the three bays (Fig. 5, r2 = 0.60). While we hope future samples will further clarify this mechanism, these data support the food quality hypothesis.
Figure 5. Relationship between diatom dietary marker and carapace width of C3 crabs from 3 Kodiak nursery embayments.
To date, our work has shown that juvenile Tanner crabs rearing in Womens Bay achieve a size advantage over juveniles rearing in other nearby embayments. Further, our data suggest this is a result of differential growth; more specifically, that there is better quality of food in Womens Bay. This size advantage may enhance crab survival as they move out of nurseries and into deeper water, where predation may be more intense. Further, more rapid growth will likely translate into earlier entry into the reproductive population and commercial population. In 2012 we will test additional hypotheses related to juvenile Tanner crab growth in Kodiak nursery embayments, with the goal of eventually expanding this work to the greater Gulf of Alaska and the Bering Sea. First we will examine the dietary issue. We suspect that the lower wave energy in Womens Bay may allow for greater retention of organic material in the form of a sediment surface “fluff” layer or benthic diatoms, which provide a superior food source for juvenile crabs. We will sample the newly settled organic material or fluff layer in Womens Bay and Pillar Creek Cove during July 2012 and analyze the material for total organic matter per dry weight, lipid classes, and fatty acid biomarkers. Crabs will also be sampled from each site and analyzed for lipids, fatty acid biomarker, and δ13C and δ15N analyses. These approaches will clarify differences in sources of organic material and elucidate whether crabs are feeding at different trophic levels in Womens and Pillar embayments. This will be accompanied by laboratory experiments, examining the effects of lipid profiles upon Tanner crab growth. Gel food diets will be formulated to mimic the differences that we have observed in essential fatty acids in the field. At the end of the experiment, in addition to quantifying crab growth, we will assess the effect of variable diet on the lipid condition of crabs. This data will be merged with our temperature growth data to develop a predictive model of juvenile Tanner growth as a function of dietary quality and temperature. Lastly, we will examine whether there is an alternative shallow water habitat (<5m) that may be accessible to settling Tanner crabs during May in Womens Bay, with periodic temperature stratification (low wave action = greater potential for warm shallows) giving crabs a temperature growth bump. These approaches should provide us with a more thorough understanding of how habitat characteristics determine nursery quality, and in turn, the function of these nurseries in supporting Tanner crab populations.
By Clifford Ryer and Louise Copeman (CIMRS, Oregon State University)