Energetic Cost of Ichthyophonus Infection in Juvenile Pacific Herring
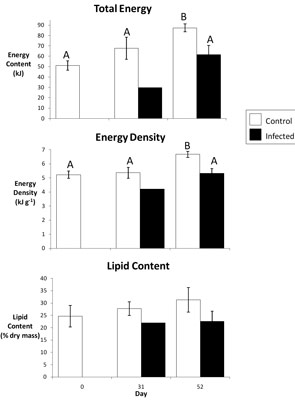
Figure 1. Energetic cost of Ichthyophonus infection in "autumn" young-of-the-year Pacific herring depicted by total energy content, energy density, and lipid content (% dry mass). Fish represented in this figure were cultured in ambient water temperature (9.5°). Different letters represent statistical differentiation. Lack of letters indicates no statistical differentiation. Low sample size of infected fish on day 31 precludes statistical tests.
Ichthyophonus is a commonly occurring parasite that has been reported in more than a hundred species of fish. It is highly pathogenic to Pacific herring, and occurs in high prevalence and intensity in herring populations throughout the northeast Pacific Ocean. Outbreaks of the parasite are known to structure herring populations.
The energetic costs of fasting and Ichthyophonus infection were measured in juvenile Pacific herring in a lab setting at three temperatures. Infected herring incurred significant energetic costs, the magnitude of which depended on fish condition at the time of infection (fat versus lean).
Herring that were fed continually and were in relatively good condition at the time of infection (fat) never stored lipid despite ad libitum (free) feeding. In feeding herring, the energetic cost of infection was a 30% reduction in total energy content relative to controls 52 days post infection (Fig. 1). Following food deprivation (lean condition), infection caused an initial delay in the compensatory response of herring.
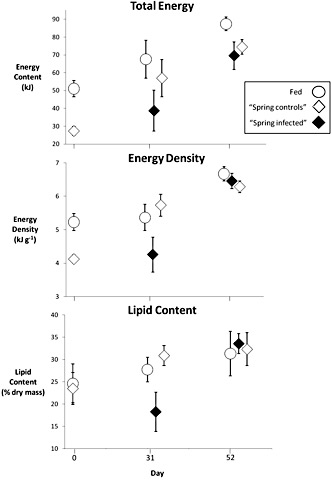
Thirty-one days after re-feeding, the energetic cost of infection in previously-fasted fish was a 32% reduction in total energy content relative to controls (Fig. 2). Body composition of infected herring subsequently recovered to some degree, though infected herring never attained the same energy content as their continuously fed counterparts.
Figure 2. Compensatory response of young-of-the-year Pacific herring infected with Ichthyophonus depicted by total energy content, energy density, and lipid content (% dry mass). Fish represented in this figure were cultured in ambient water temperature (9.5°). Low sample size of infected fish on day 31 precludes statistical tests.
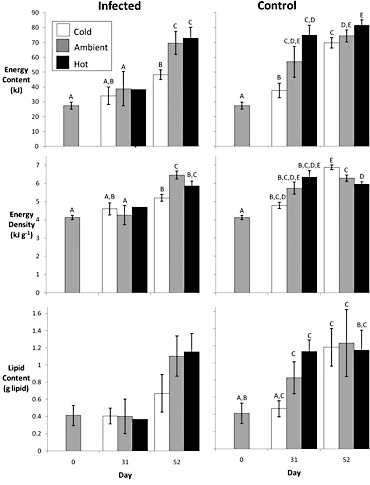
Figure 3. Temperature influence on the energetic cost of Ichthyophonus infection in "spring" young-of-the-year Pacific herring depicted by total energy content, energy density, and lipid content (g lipid). Water temperatures were cold (9.5°), ambient (12.0°), and hot (15.0°). Different letters within a panel represent statistical differentiation. Lack of letters indicates no statistical differentiation. Low sample size of infected fish on day 31 precludes statistical tests.
Fifty-two days after re-feeding, the energetic cost of infection in previously-fasted fish was a 6% reduction in total energy content relative to controls. The greatest impacts of infection occurred in colder temperatures, suggesting Ichthyophonus- induced reductions in body condition may have greater consequences in the northern extent of herring's range, where juveniles use most of their energy reserves to survive their first winter (Fig. 3).